Mastrycht
| System | Oddział | Piętro | Wiek (mln lat) |
|---|---|---|---|
| Paleogen | Paleocen | Dan | młodsze |
| Kreda | Górna | Mastrycht | 66,0–72,1 |
| Kampan | 72,1–83,6 | ||
| Santon | 83,6–86,3 | ||
| Koniak | 86,3–89,8 | ||
| Turon | 89,8–93,9 | ||
| Cenoman | 93,9–100,5 | ||
| Dolna | Alb | 100,5–113,0 | |
| Apt | 113,0–125,0 | ||
| Barrem | 125,0–129,4 | ||
| Hoteryw | 129,4–132,9 | ||
| Walanżyn | 132,9–139,8 | ||
| Berrias | 139,8–145,0 | ||
| Jura | Górna | Tyton | starsze |
| Podział według IUGS, luty 2017 | |||
Mastrycht (ang. Maastrichtian)
- w sensie geochronologicznym – szósty, najmłodszy, wiek późnej kredy, trwający około 6,1 milionów lat (od 72,1 ± 0,2 do 66,0 mln lat temu). Mastrycht jest młodszy od kampanu a starszy od danu (paleocen).
- w sensie chronostratygraficznym – szóste, najwyższe, piętro górnej kredy, wyższe od kampanu a niższe od danu.
Stratotyp dolnej granicy mastrychtu znajduje się w Tercis les Bains nad rzeką Adour (Landes, Francja). Granica opiera się na pierwszym wystąpieniu amonita Pachydiscus neubergicus (Hauer, 1858).
Nazwa pochodzi od miasta Maastricht w Limburgii (Holandia).
Był to ostatni okres, w którym żyły nieptasie dinozaury, pterozaury, plezjozaury i mozazaury.
Fauna kręgowa
| Ssaki mastrychtu | ||||
|---|---|---|---|---|
| Takson | Występowanie | Miejsce | Opis | Grafika |
|
Montana | Jeden z największych mezozoicznych ssaków. | 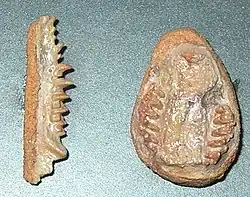 | |
| Montana | Uważany za najwcześniejszego przedstawiciela naczelnych lub Primatomorpha | |||
| Mongolia | ||||
| Ptaki mastrychtu | ||||
|---|---|---|---|---|
| Takson | Występowanie | Miejsce | Opis | Grafika |
|
Rodzaj ptaków spokrewnionych ze współczesnymi kaczkami i gęsiami. Znaleziony w formacji Hornerstone w New Jersey. Starszy z dwóch znanych gatunków Anatalavis. Młodszy, z wczesnego eocenu Londynu, nosi nazwę A. oxfordi. |  .jpg) | ||
|
Rodzaj drapieżnego ptaka. Obejmuje dwa gatunki, oba występujące w mastrychcie. Gatunek typowy, Avisaurus archibaldi, został odkryty w formacji Hell Creek w stanie Montana. | |||
|
Monotypowy rodzaj nielotnego uzębionego morskiego ptaka, spokrewniony z hesperornisem. Był to duży ptak (ponad 1,5 m długości) żyjący w płytkich wodach na terenach dzisiejszej północnej Kanady. | |||
|
Przedstawiciel siewkowych z formacji Lance Creek. Jego relacje ewolucyjne są niejasne. Znany jedynie z niekompletnej kości kruczej C. major jest jedynym znanym gatunkiem z rodzaju Ceramornis. | |||
|
Rodzaj siewkowatych z formacji Lance Creek w stanie Wyoming. | |||
|
Rodzaj ptaka drapieżnego odkryty w Argentynie. Prawdopodobnie spokrewniony z Avisaurus. Metrowej długości przypominający orła drapieżca, jedyny znany gatunek rodzaju to Enantiornis leali. | |||
|
Rodzaj dużych nielotów podobnych do bezgrzebieniowych. Jaja przypisywane wcześniej tytanozaurom mogą w rzeczywistości należeć do przedstawicieli Gargantuavis. Znany z terenów dzisiejszej Francji Gargantuavis philoinos jest jedynym znanym gatunkiem rodzaju. | |||
|
||||
| Duży wodny nurkujący ptak z zębami, szczątkowymi skrzydłami i płatowatymi palcami. Odkryty w Kansas przez Marsha podczas wojny o kości. | ||||
|
Stosunkowo prymitywny rodzaj spokrewniony z hesperornisem. Ten uzębiony nielotny ptak żył w wodach słodkich, w przeciwieństwie do swych krewnych. Jedyny znany gatunek rodzaju to Judinornis nogontsavensis. Żył w estuariach i rzekach gór formacji Nemegt w Mongolii. | |||
|
||||
|
Rodzaj ptaka brodzącego o niepewnej pozycji systematycznej, przypominał wielkością i budową nóg współczesnego kulika. Jedyny znany gatunek, żył w Argentynie | |||
|
Ptak morski z Chile. Miał śródstopie ptaka nurkującego z pomocą nóg, ale jego pokrewieństwo jest nieznane. Jedyny znany gatunek żył na granicy kampanu i mastrychtu. | |||
|
||||
|
||||
|
Rodzaj ptaków z formacji Lance Creek spokrewniony z hesperornisem.
Typowy (i jedyny znany gatunek) otrzymał epitet na cześć doktora Alexandra F. Skutcha. Odkryte szczątki obejmują tylko prawą kość kwadratową i fragmenty szkieletu pozaczaszkowego. | |||
|
||||
|
||||
|
Krewny współczesnych kaczek i gęsi. Jedyny znany gatunek rodzaju, Vegavis iaai, jest znany ze skamieniałości jednego osobnika odkrytych na Antarktydzie. | |||
†Teropody (nie-ptasie)
| †Nie-ptasie teropody mastrychtu | ||||
|---|---|---|---|---|
| Takson | Występowanie | Miejsce | Opis | Grafika |
|
Formacja Allen?, Formacja Anacleto?, Argentyna | Dwunożny drapieżnik osiągający prawdopodobnie 7–9 m długości; znany jedynie z jednej niekompletnej czaszki |  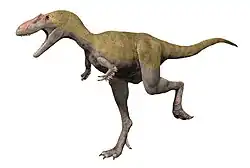  .png)   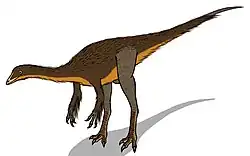      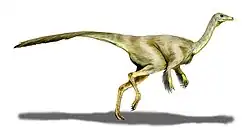     | |
|
Formacja Nemegt, Mongolia | Dromeozauryd, osiągający prawdopodobnie około 2,5 m długości, wyróżniający się stosunkowo małymi pazurami na drugich palcach stóp. | ||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
Słabo poznany rodzaj mierzącego około 1,8 m drapieżnika z rodziny troodontów. Jego szczątki odnaleziono w Mongolii. Zawdzięcza swą nazwę „borogom” (ang. „borogoves”) z wiersza Jabberwocky Lewisa Carrolla. Znany jedynie z niekompletnych kończyn przednich. | |||
|
Dawniej uważany za wielką sowę. Jego nazwa znaczy „zły wolnostop”. Znaleziony w Transylwanii, aluzja do hrabiego Drakuli może być zamierzona. | |||
|
Duży, mierzący około 10 m długości drapieżnik, żyjący na terenie dzisiejszej Argentyny. Carnotaurus cechował się krótkim pyskiem, niewielkimi różkami nad oczami i bardzo krótkimi kończynami przednimi. Znany z jednego kompletnego szkieletu oraz odcisków skóry dowodzących, że nie był opierzony. | |||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
Może być młodym osobnikiem Tyrannosaurus lub innego rodzaju Tyrannosauridae[1]. | |||
|
||||
|
||||
|
||||
|
||||
| Formacja Nanxiong, Chiny | ||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
| †Zauropody mastrychtu | ||||
|---|---|---|---|---|
| Takson | Występowanie | Miejsce | Opis | Grafika |
|
   | |||
|
||||
| Bruhathkayosaurus mógł być największym dinozaurem. | ||||
|
||||
|
Grès à Reptiles, Francja; Hiszpania | |||
|
Indie | |||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
| †Ceratopsy mastrychtu | ||||
|---|---|---|---|---|
| Takson | Występowanie | Miejsce | Opis | Grafika |
|
|
         | ||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
USA | |||
| †Pachycefalozaury mastrychtu | ||||
|---|---|---|---|---|
| Takson | Występowanie | Miejsce | Opis | Grafika |
|
 .jpg)  | |||
|
||||
|
||||
|
||||
|
||||
| †Ornitopody mastrychtu | ||||
|---|---|---|---|---|
| Takson | Występowanie | Miejsce | Opis | Grafika |
|
            | |||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
| †Ankylozaury mastrychtu | ||||
|---|---|---|---|---|
| Takson | Występowanie | Miejsce | Opis | Grafika |
|
Potwierdzone | Największy ankylozaur |    | |
| Kampan do mastrychtu | Formacja Horseshoe Canyon, Alberta, Kanada | Masywny nodozaur mierzący około 6,6 m długości i 2 m wysokości. Miał małe, nierówne kostne płytki na grzbiecie i głowie oraz ostre kolce na grzbiecie i ogonie. Cztery największe kolce wyrastały z ramion po każdej stronie, dwa z nich dzieliły się u niektórych gatunków na części. Widziana z góry czaszka miała kształt gruszki. | ||
|
||||
|
||||
|
||||
| †Pterozaury mastrychtu | ||||
|---|---|---|---|---|
| Takson | Występowanie | Miejsce | Opis | Grafika |
|
Formacja Densus-Ciula, Rumunia, jeden z największych znanych pterozaurów (rozpiętość skrzydeł ok. 12 m) |   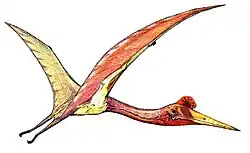 | ||
|
Ameryka Północna | |||
|
Ameryka Północna, jeden z największych znanych pterozaurów (rozpiętość skrzydeł ok. 10 m) | |||
| Krokodyle mastrychtu | ||||
|---|---|---|---|---|
| Takson | Występowanie | Miejsce | Opis | Grafika |
 | ||||
| †Plezjozaury mastrychtu | ||||
|---|---|---|---|---|
| Takson | Występowanie | Miejsce | Opis | Grafika |
| Nowa Zelandia | Formacja Conway | elasmosaur | ||
|
 | |||
| Angola | Formacja Mocuio | |||
|
||||
| Łuskonośne mastrychtu | ||||
|---|---|---|---|---|
| Takson | Występowanie | Miejsce | Opis | Grafika |
|
Mozazaur |    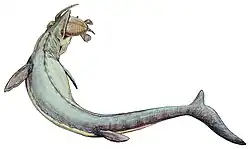 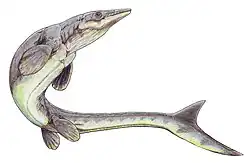     | ||
| Mozazaur | ||||
|
Mozazaur | |||
|
Mozazaur | |||
|
Mozazaur | |||
|
Mozazaur | |||
|
Mozazaur | |||
|
Mozazaur | |||
|
Wąż | |||
|
Mozazaur | |||
|
Mozazaur | |||
|
Mozazaur | |||
|
Mozazaur | |||
|
Mozazaur | |||
|
Mozazaur | |||
| Mozazaur | ||||
| Żółwie mastrychtu | ||||
|---|---|---|---|---|
| Takson | Występowanie | Miejsce | Opis | Grafika |
|
||||
|
||||
|
Gatunek żółwia morskiego z Kansas uznany mylnie za hadrozaura przez E.D. Cope’a. Jedyny znany obecnie gatunek Pneumatoarthrus. | |||
| Płazy mastrychtu | ||||
|---|---|---|---|---|
| Takson | Występowanie | Miejsce | Opis | Grafika |
|
|
 | ||
|
|
 | ||
| Ryby kostnoszkieletowe mastrychtu | ||||
|---|---|---|---|---|
| Takson | Występowanie | Miejsce | Opis | Grafika |
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
| Ryby chrzęstnoszkieletowe mastrychtu | ||||
|---|---|---|---|---|
| Takson | Występowanie | Miejsce | Opis | Grafika |
|
 | |||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
|
||||
Flora
Zobacz też
Przypisy
- ↑ Henderson (2005). „Nano No More: The death of the pygmy tyrant.” In „The origin, systematics, and paleobiology of Tyrannosauridae”, a symposium hosted jointly by Burpee Museum of Natural History and Northern Illinois University.