Ostrogony
| Xiphosura[1] | |
| Latreille, 1802 | |
_001.jpeg) Tachypleus tridentatus | |
| Systematyka | |
| Domena | |
|---|---|
| Królestwo | |
| Podkrólestwo | |
| Infrakrólestwo | |
| (bez rangi) | pierwouste |
| Nadtyp | |
| (bez rangi) | Panarthropoda |
| Typ | |
| Podtyp | |
| Nadgromada | |
| Gromada |
ostrogony |
Ostrogony (Xiphosura) – gromada morskich, wszystkożernych stawonogów z grupy szczękoczułkowców właściwych (Euchelicerata). Dawniej uznawane były razem z wymarłymi wielkorakami i skorpionami jako rząd wliczany do gromady staroraków (Merostomata). Współcześnie żyją 4 gatunki skupione w rodzinie skrzypłoczy (Limulidae). Prosoma o podkowiastym, kanciastym kształcie, z zestawem oczu prostych i złożonych. Opistosoma jest zakończona długim i ostrym kolcem, który działa jak ster. Odnóża znajdują się pod pancerzem, pomiędzy którym jest otwór gębowy. Rozmiarami dochodzą do 60, maks. 85 cm długości. W okresie rozrodczym gromadzą się przy brzegach wód i strefie pływów na lądzie.
Najbardziej znanym przedstawicielem jest skrzypłocz Limulus polyphemus, zamieszkujący wybrzeże Atlantyku w Ameryce Północnej.
Terminologia
W języku polskim ostrogony nazywane są także skrzypłoczami[2] albo mieczogonami[3].
Charakterystyka
_adult_underside_(26923436154).jpg)
Wygląd zewnętrzny
_(6707310189).jpg)
Ostrogony to duże szczękoczułkopodobne, osiągające 60, maksymalnie 85 cm długości; największy gatunek to Tachypleus gigas, jeden z 2 taksonów swego rodzaju[4]. Mają grubą kutikulę (o grubości 3 mm) pozbawioną wapiennych wtrętów, o barwie ciemnobrunatnej lub czarnobrązowej. We wnętrzu ciała znajduje się mezodermalny endoszkielet. Samice większe od samców[4].
Młode osobniki z wyglądu przypominają wymarłe trylobity (Trilobita), natomiast formy dorosłe bywają brane za przedstawicieli skorupiaków (Crustacea). Ciało ostrogonów podzielone jest na sześciokątną, podkowiastą prosomę okoloną karapaksem oraz opistosomę, na którą z kolei składa się mezosoma oraz metasoma[4]. Złożona z 6 segmentów prosoma nie wykazuje śladów członowania na stronie grzbietowej, na bokach jej karapaksu znajdują się oczy złożone o nerkowatym kształcie, zaś z przodu występują proste oczy środkowe[4]. Mezosoma jest zbudowana z 7 segmentów (widoczne to u młodych osobników, słabo zaznaczone u dorosłych), zaopatrzona w pleurostergity, czyli boczne płytki posiadające ruchome kolce[4]. Metasoma ma jeden długi kolec, zwany tergalnym, który powstał wskutek zrośnięcia się telsonu i kilku segmentów opistosomy[4]. Służy jako ster w trakcie pływania, a także zapewnia zachować balans ciała[5], ostrogony nie używają go w celu kłucia. Na lądzie z kolei zwierzę może odwrócić się na grzbiet po upadku na tę stronę ciała właśnie dzięki kolcowi[4]. Staw między przednią a tylną częścią ciała umożliwia zginanie się na brzuszną stronę, ale nie jest on odpowiednikiem granicy między prosomą a opistosomą spotykaną u pajęczaków[4].
.jpg)
Pierwsza para odnóży, które są ułożone na prosomie, to trójczłonowe szczękoczułki (chelicery), które zakończone są małymi szczypcami; znajdują się przy otworze gębowym. Odnóża kroczne pełniące funkcje lokomotoryczne są znacznie większe od chelicer, mieszczą się na spodniej stronie karapaksu w liczbie 5 par[4]. Na biodrach, członach podstawowych, tych odnóży osadzone są endyty – wyrostki żujące posiadające szczeciny; 4 pary spośród tych odnóży są zakończone szczypcami, podczas gdy 5. para zaopatrzona jest w szczeciny (służą do niezapadania się w grząskim podłożu zwierzęcia) oraz flabella (nakierowują prąd wody w stronę jamy skrzelowej, zapobiegają dostawaniu się zanieczyszczeń, mogą pełnić funkcje zmysłowe)[4]. Odnóża mezosomy uległy modyfikacji; pierwsza para kończyn jest zredukowana i tworzy małe wyrostki znane jako chilaria. Pozostałe 6 par są dwugałęziste, przybierają formę płytek, w ramach każdej pary są one zrośnięte; pierwsza para jest najgrubsza i przykrywa pozostałe odnóża tworząc wieczko (operculum), reszta tych odnóży przekształconych jest w skrzela[4]. Metasoma nie ma odnóży[4].
Budowa wewnętrzna

Układ nerwowy i percepcja
Mózg, ponieważ otwór gębowy uległ przemieszczeniu na brzuszną stronę ciała, nie leży nad przełykiem[4]. Nerwy, które są przednimi odnogami mózgu tworzą unerwienie oczu. Ponad ¾ masy całego mózgu tworzą ciałka grzybkowate (corpora pedunculata), które są ośrodkami kojarzeniowymi protocerebrum. Tritocerebrum jest wyzbyte komisury podprzełykowej[4]. Oprócz neuromerów i nerwów odpowiedzialnych za unerwienie odnóży prosomy, w skład zwoju podprzełykowego wchodzą neuromery pierwszego i drugiego segmentu opistosomy. Segmenty odnóży skrzelowych unerwiają zwoje i nerwy ulokowane poza obszarem mózgu na parzystym, brzusznym pniu nerwowym[4]. Całe ciało okryte jest przez zmysłowe szczeciny dotykowe a także szczeciny o nieznane jak dotąd funkcji umieszczone w zagłębieniach[4].
Za narządy zmysłów służą nie tylko organy zmysłowe, ale i oczy[4]. Fasetki (omatidia), z których to zbudowane są boczne oczy złożone ostrogonów, są prościej zbudowane niż u tych zauważalnych u żuwaczkowców (Mandibulata), gdyż nie posiadają stożka krystalicznego; soczewka rogówki, jaką jest kutykula okrywająca całe oczy złożone, tworzy wypustki skupiające źródło światła w poszczególnych fasetkach[4]. Rabdom należący do pojedynczej fasetki ma pałeczkowaty kształt, jest zbudowany z 8–20 komórek siatkówki, mając na przekroju poprzecznym kształt gwiazdy; wyrostek bocznie leżącej komórki świetlnej wnika do wnętrza rabdomu[4]. Aksony komórek zmysłowych sąsiadujących fasetek łączą się ze sobą; za zwiększenie kontrastu widzenia odpowiadają aksony, które działają hamująco na omatidia. Z kolei fasetki ulokowane w tylnej części oka są zredukowane i możliwe, że pełnią funkcję neurosekrecyjną[4].

Układ oddechowy
Począwszy od 3, kończąc na 7 segmencie mezosomy, ujrzeć można pięć par odnóży oddechowych (skrzela)[4]. Każda zewnętrzna gałąź tych odnóży posiada do 150 blaszek układających jedna na drugiej na wzór kartek w książce – są one ukrwione i obmywane przez wodę. Telepodity tych odnóży przypominają małe, trójczłonowane wyrostki, a ich zadaniem jest oczyszczanie skrzeli[4]. Pajęczaki, takie jak zdecydowana większość pająków (Araneae) skorpiony (Scorpiones), biczykoodwłokowce (Thelyphonida), rozłupnogłowce (Schizonomida) oraz tępoodwłokowce (Amblypygi) oddychają za pomocą blaszkowatych płuc książkowych, które są ewolucyjną pozostałością po wodnych przodkach i są to homologi względem narządów oddechowych ostrogonów[6][7].
Układ wydalniczy
Gruczoły biodrowe (koksalne) spełniają rolę narządów wydalniczych.
Lokomocja
W czasie chodzenia po twardym podłożu wykorzystywane są odnóża prosomy, poruszają się one naprzemiennie[4]. Kiedy ostrogon pływa, synchronicznie wiosłuje odnóżami prosomy a także mezosomy. Zaostrzona krawędź prosomy w kontakcie z miękkim, mulistym lub piaszczystym podłożem działa na wzór pługu, co umożliwia ukrycie się przed drapieżnikiem na skutek zagrzebania[4]. Jak wspomniano, w lokomocji rolę odgrywa kolec tergalny zwierzęcia.
Występowanie
Rodzaj Limulus (1 gatunek) występuje u wybrzeży Atlantyku (od amerykańskich stanów Nowy Jork i Florydy sięgając do Jukatanu), rodzaj Carcinoscorpius (1 gatunek) spotykany jest w Zatoce Bengalskiej, u wód Malezji i Filipin, natomiast Tachypleus (2 gatunki) notowany jest z Azji Południowej[4]. Ostrogony stwierdzono także u wybrzeży Japonii.
Ekologia
Zwierzęta te zamieszkują płytkie i ciepłe morza, ich szelfy, do głębokości 10–40 m[4]. Zagrzebują się w podłożu, w mule i tam polują na wolno poruszającą się zdobycz – w uogólnieniu, w składzie ich diety znajdują się drobne skorupiaki (Crustacea), pierścienice (Annelida) i małże (Bivalvia), chociaż spożywają również padlinę[4]. Dla przykładu, Tachypleus gigas został opisany jako stworzenie bentosu, odżywiające się mięczakami, wieloszczetami albo rozkładającą się materią organiczną[8]. W diecie młodych ostrogonów występują glony; badania jednocześnie pokazały, że młode karmione w >70% białkiem zwiększały swój rozmiar, ale jednocześnie potomstwo wykazywało większą śmiertelność w porównaniu do tego, które karmiono dietą z glonów[9]. Nocą lub w okresie rozrodczym (zob.: cykl życiowy) ostrogony wędrują po suchym lądzie przy brzegu[4].
Na kuli ziemskiej ostrogony są rozmieszczone nieregularnie; ich dysjunktywne rozmieszczenie może wynikać z długiej historii ewolucyjnej, na którą prawdopodobnie wywarła piętno historia geologiczna Ziemi (zmiany klimatyczne, podział kontynentów)[4]. Kopalne Xiphosura w porównaniu do dzisiejszych przedstawicieli gromady były kosmopolityczne, a współczesne taksony to pozostałość po tej bioróżnorodności[4].
Makroskamieniałości przynależące do ostrogonów znajdywane w osadach skalnych są określane jako rzadkie, jedne z rzadszych[10].
Rozmnażanie i cykl życiowy
.jpg)
_003.jpeg)
Okres godów przypada na wczesne lato kiedy to omawiane stawonogi gromadzą się w strefie pływów albo płaskich brzegach mórz[4]. Jak wspomniano, samice są większe od samców. U samców Limulus szczypce pedipalp, odnóży 1. pary, są powiększone i pełnią funkcje chwytne; w podobny sposób u dwóch kolejnych rodzajów są poddane modyfikacjom szczypce 2. pary odnóży[4]. Samce dzięki takim modyfikacjom mogą przytrzymywać swe partnerki i pozostawać przyczepione do ich ciał nawet tydzień zanim dojdzie między osobnikami do kopulacji.
Samice w płytkich nieckach składają od 200 do 1000 jaj, a samce polewają je nasieniem, dołek po tym akcie następnie zostaje zasypany piaskiem[4]. W ciągu całego sezonu rozrodczego, samice ostrogonów mogą złożyć w takich dołkach od 60 do 120 0000 jaj[4]. W przypadku Carcinoscorpius rotundicauda, akt kopulacji a także składanie jaj następuje w wodach słodkich i słonawych, a następnie samica nosi zapłodnione jaja na swych odnóżach mezosomy[4].
Bruzdkowanie jest całkowite i nierównomierne, a gastrulacja odbywa się poprzez imigrację[4]. Po około 2 tygodniach od złożenia jaj wykluwają się z nich larwy noszące nazwę larw trylobitowych, które odżywiają się zapasami żółtka. W przypadku jednego z badań przeprowadzonego w warunkach laboratoryjnych młode wykluły się po 6 tygodniach[11]. Larwa trylobitowa ma dobrze ukształtowaną segmentację, ale 9 par odnóży[4]. Kolejnym stadium rozwojowym jest presterichia, która ma już wykształcony kolec tergalny, a także pełen zestaw odnóży – presterichia to określenie na ostrogona, który przeszedł pierwsze linienie postembrionalne[4]. W ciągu pierwszego roku swego życia larwa przechodzi przez 6 kolejnych linień, podczas gdy osobniki dorosłe wykonują tą czynność raz do roku[4].
Xiphosura dojrzewają płciowo w wieku 9–12 lat, a gatunek Limulus polyphemus dożywa 14–19 lat[4].
Taksonomia
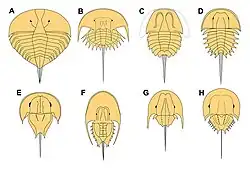
Historia badań
Ostrogony przynależą do grupy szczękoczułkopodobnych (Cheliceromorpha) i są wymieniane przez badaczy jako zwierzęta archaiczne, żywe skamieniałości[12]. Takson Xiphosura odznacza się tym, że jest to punktualiczna linia rozwojowa[12]. W przeszłości uznawano je za grupę siostrzaną wobec wymarłych wielkoraków (Eurypterida), wliczając obie te grupy do wspólnej gromady staroraków (Merostomata)[4]. Według tego poglądu nie byłyby jedynymi przedstawicielami tej grupy: skorpiony (Scorpiones) z powodu swego podobieństwa w morfologii do Eurypterida również zaliczane były do staroraków, jednocześnie były one sympatryczne w stosunku do tych drugich (już) od czasów syluru (450 mln lat)[13]. Merostomata, zawierając te 3 grupy, miałaby jakoby być taksonem siostrzanym do pajęczaków (Arachnida)[4]. Jednakże, badania oparte na filogenezie oraz materiale kopalnym nie wspierają istnienia grupy, jaką są staroraki, jest to wedle danych jednostka parafiletyczna[4]. Wykazano dodatkowo, że wymarłe wielkoraki są bliżej spokrewnione z pajęczakami, do których wliczono ostatecznie też skorpiony[14]; konsekwencją tych rozważań filogenetycznych okazało się wskrzeszenie taksonu znanego jako szczękoczułkowce właściwe (Euchelicerata), zrzeszającego ostrogony, wielkoraki i pajęczaki[4]. Euchelicerata zostały ulokowane w obrębie szczękoczułkopodobnych, będąc grupą na tym samym poziomie, co kikutnice (Pycnogonida) i nieżyjące już Aglaspida[a] – wszystkie klady są nadgromadami[15].
Kwestia odrębności Xiphosura
Badania przeprowadzone przez Lamsdella (2013) ujawniają, że ostrogony, podobnie jak grupa staroraki, też niekoniecznie są monofiletyczne[16]. W opracowaniu Dunlopa, Penney'a oraz Jekel (stan na rok 2018) poświęconemu wymarłym pajęczakom i ich krewnym przy nazwie Xiphosura dopisano zwrot sensu lato[17]. W przypadku samych pajęczaków, w niektórych badaniach są również wyłaniane jako grupa odznaczająca się parafiletyzmem[18]. W związku z tym, Ballesteros i Sharma (2019) stwierdzili, że ostrogony powinne zostać umieszczone w gromadzie pajęczaków, ażeby uzyskać monofiletyzm Arachnida. Hipoteza ta była powtarzana przez Ballesterosa i innych w kolejnych latach[19][20]. Najbliższymi krewnymi Xiphosura okazywałyby się wtedy kapturce (Ricinulei)[18] – rząd pajęczaków, który pierwotnie był uważany raczej za krewnych roztoczy (Acari)[21]. Zgodnie z tą teorią, Xiphosura nie powinny być traktowane jako siostrzane w stosunku do Arachnida, a same w sobie być uznawane za ich wodnych przedstawicieli, podobnie jak jest to w przypadku wodopójek („Hydrachnidia”, też wodne)[18]. W odpowiedzi na analizę Ballesterosa i Sharma'y, zespół badaczy pod przewodnictwem Fernandeza w tym samym roku zaprzeczył hipotezie tych autorów – według nich, kikutnice oraz ostrogony wciąż należy uznawać za siostrzane do pajęczaków, zaś najbliższymi krewnymi kapturców są kosarze (Opiliones); jednocześnie, według badaczy Arachnida są monofiletyczne, jest to ugruntowane[22].
Ewolucja i systematyka

Ewolucja
Mówi się, że ostrogony jako grupa ukształtowała się w ordowiku[12], ale znana jest ona już od syluru, a niektóre dane wskazują, że jej początki sięgają kambru (500 mln lat temu)[4]. Jako żywe skamieniałości, ich anatomia w ogóle lub nieznacznie się zmieniła na przestrzeni lat ewolucji[12]. W przeszłości nie tylko występowały w morzach, ale poszczególne taksony Xiphosura wielokrotnie i niezależnie opanowały również środowiska słodkowodne[12]. Współcześnie występujące ostrogony różnią się nieznacznie od wymarłych krewnych, takich jak np. Mesolimulus[4].
Podział systematyczny

Obecnie wyróżnia się 104 taksony tych zwierząt[17], z czego 4 to żyjące gatunki, skupione w jednej rodzinie; znane także są liczne taksony, które żyły w paleozoiku oraz mezozoiku[12]. Obecnie żyjąca rodzina Limulidae, znana jako skrzypłocze istnieje od triasu[4]. System internetowy World Register of Marine Species (WoRMS) dodatkowo dzieli rodzinę Limulidae na dwie podrodziny: Limulinae i Tachypleinae, terminy wykreowane kolejno przez Williama Elforda Leacha (1819) i Reginalda Innesa Pococka (1902). Badania ukazują, że Limulus jest siostrzany wobec Carcinoscorpius i Tachypleus[10]. Poniżej zaprezentowano podział systematyczny współczesnych Limulidae opierający się na tym systemie[23]:
- Limulinae Leach, 1819
- Limulus O.F. Mueller, 1785
- Limulus polyphemus Linnaeus, 1758
- Limulus O.F. Mueller, 1785
- Tachypleinae Pocock, 1902
- Carcinoscorpius Pocock, 1902
- Carcinoscorpius rotundicauda Latreille, 1802
- Tachypleus Leach, 1819
- Tachypleus gigas O.F. Mueller, 1785
- Tachypleus tridentatus Leach, 1819
- Carcinoscorpius Pocock, 1902
oraz wymarłe gatunki z rodzaju:
- †Mesolimulus
Pozostałe, wymarłe już gatunki klasyfikowane były w podrzędzie †Synziphosurida. Jednakże, jak wspomniano, Lamsdell zauważył, że Xiphosura nie są monofiletyczne, a Synziphosurida nie są grupą pnia ostrogonów, tylko szczękoczułkowców. W kolejnych pracach próbowano odzwierciedlić to stanowisko, lecz w tym schemacie klad Planaterga obejmowałby również Chasmataspidida, Eurypterida i Arachnida, a Planaterga jest zagnieżdżona w Prosomapoda[17].
Znaczenie gospodarcze
.jpg)
Znaczenie ostrogonów w życiu człowieka określane jest jako niewielkie[4].
Przede wszystkim, w krajach Orientu skrzypłocze są używane w celach kulinarnych[4]. Dla przykładu, w Tajlandii te zwierzęta nie są chronione, a ich liczebność ich populacji poważnie spada. Jednakże, spożywanie skrzypłoczy może poskutkować zatruciem wywołanym tetrodotoksyną – mięso ze skrzypłocza Carcinoscorpius rotundicauda.
Uwagi
- ↑ Kwestia spokrewnienia Aglaspida ze współcześnie żyjącymi Arthropoda jest sporna. Jedni zaliczają je jako bliskie szczękoczułkowcom, a inni – żuwaczkowcom.
Przypisy
- ↑ Xiphosura, [w:] Integrated Taxonomic Information System (ang.).
- ↑ skrzypłocz, [w:] Internetowa Encyklopedia PWN, Wydawnictwo Naukowe PWN [dostęp 2025-03-16] (pol.).
- ↑ mieczogon, [w:] Internetowa Encyklopedia PWN, Wydawnictwo Naukowe PWN [dostęp 2025-03-16] (pol.).
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 Jacek Dabert: Gromada: ostrogony – Xiphosurida. W: Zoologia: Stawonogi. T. 2, cz. 1. Szczękoczułkopodobne, skorupiaki. Czesław Błaszak (red. nauk.). Warszawa: Wydawnictwo Naukowe PWN, 2011. ISBN 978-83-01-17447-7.
- ↑ Gerald E. Silvey, Motor Control of Tail Spine Rotation of the Horseshoe Crab, Limulus Polyphemus, „Journal of Experimental Biology”, 3, 58, 1973, s. 599–625, DOI: 10.1242/jeb.58.3.599 [dostęp 2025-03-16] (ang.).
- ↑ Czesław Błaszak: Gromada: pajęczaki – Arachnida. W: Zoologia: Stawonogi. T. 2, cz. 1. Szczękoczułkopodobne, skorupiaki. Czesław Błaszak (red. nauk.). Warszawa: Wydawnictwo Naukowe PWN, 2011, s. 51-63. ISBN 978-83-01-17447-7.
- ↑ Czesław Jura: Bezkręgowce. Podstawy morfologii funkcjonalne, systematyki i filogenezy. Wyd. 3. Warszawa: PWN, 2005, s. 488-531.
- ↑ Anil Chatterji, J.K. Mishra & A.H. Parulekar, Feeding behaviour and food selection in the horseshoe crab, Tachypleus gigas (Müller), „Hydrobiologia”, 246, 1992, s. 41–48, DOI: 10.1007/BF00005621 [dostęp 2025-03-16] (ang.).
- ↑ Diet Composition of Juvenile Horseshoe Crabs: Implications for Growth and Survival of Natural and Cultured Stocks, [w:] Ruth H. Carmichael i inni, Biology and Conservation of Horseshoe Crabs, Springer, 2009, s. 521–534, DOI: 10.1007/978-0-387-89959-6_33 [dostęp 2025-03-16] (ang.).
- 1 2 Adrian Kin, Błażej Błażejowski, The Horseshoe Crab of the Genus Limulus: Living Fossil or Stabilomorph?, „PLoS ONE”, 2014, DOI: 10.1371/journal.pone.0108036 [dostęp 2025-03-16] (ang.).
- ↑ Steffen Harzsch i inni, Immunohistochemical localization of neurotransmitters in the nervous system of larval Limulus polyphemus (Chelicerata, Xiphosura): evidence for a conserved protocerebral architecture in Euarthropoda, wyd. 3, t. 34, 2005, DOI: 10.1016/j.asd.2005.01.006 [dostęp 2025-03-16] (ang.).
- 1 2 3 4 5 6 James C. Lamsdell, Horseshoe crab phylogeny and independent colonizations of fresh water: ecological invasion as a driver for morphological innovation, „Palaeontology”, 2, 59, 2015, s. 181-194, DOI: 10.1111/pala.12220 [dostęp 2025-03-15] (ang.).
- ↑ Marek Żabka: Rząd: skorpiony – Scorpionida. W: Zoologia: Stawonogi. T. 2, cz. 1. Szczękoczułkopodobne, skorupiaki. Czesław Błaszak (red. nauk.). Warszawa: Wydawnictwo Naukowe PWN, 2011. ISBN 978-83-01-17447-7.
- ↑ Richard J. Howard i inni, Exploring the evolution and terrestrialization of scorpions (Arachnida: Scorpiones) with rocks and clocks, „Organisms Diversity & Evolution”, 19, 2019, s. 71–86, DOI: 10.1007/s13127-019-00390-7 [dostęp 2025-03-15] (ang.).
- ↑ Czesław Błaszak: Podtyp: szczękoczułkopodobne – Cheliceromorpha; Nadgromada: szczękoczułkowce – Chelicerata. W: Zoologia: Stawonogi. T. 2, cz. 1. Szczękoczułkopodobne, skorupiaki. Czesław Błaszak (red. nauk.). Warszawa: Wydawnictwo Naukowe PWN, 2011. ISBN 978-83-01-17447-7.
- ↑ James C. Lamsdell, Revised systematics of Palaeozoic ‘horseshoe crabs’ and the myth of monophyletic Xiphosura, „Zoological Journal of the Linnean Society”, 1, 167, 2013, s. 1–27, DOI: 10.1111/j.1096-3642.2012.00874.x [dostęp 2025-03-16] (ang.).
- 1 2 3 Dunlop i inni, A summary list of fossil spiders and their relatives, [w:] World Spider Catalogue (WSC) [online], 2018 [dostęp 2025-03-25] (ang.).
- 1 2 3 Jesús A Ballesteros, Prashant P Sharma, A Critical Appraisal of the Placement of Xiphosura (Chelicerata) with Account of Known Sources of Phylogenetic Error, „Systematic Biology”, 6, 68, 2019, s. 896–917, DOI: 10.1093/sysbio/syz011 [dostęp 2025-03-25] (ang.).
- ↑ Jesús A Ballesteros i inni, Ordered phylogenomic subsampling enables diagnosis of systematic errors in the placement of the enigmatic arachnid order Palpigradi, „Proceedings of the Royal Society - Biological Sciences”, 2019, DOI: 10.1098/rspb.2019.2426 [dostęp 2025-03-25] (ang.).
- ↑ Jesús A Ballesteros i inni, Comprehensive Species Sampling and Sophisticated Algorithmic Approaches Refute the Monophyly of Arachnida, „Molecular Biology and Evolution”, 2, 39, 2022, s. 1-16, DOI: 10.1093/molbev/msac021 [dostęp 2025-03-25] (ang.).
- ↑ P. Weygoldt, H.F. Paulus, Untersuchungen zur Morphologie, Taxonomie und Phylogenie der Chelicerata1 II. Cladogramme und die Entfaltung der Chelicerata, „Journal of Zoological Systematics and Evolutionary Research”, 1979, DOI: 10.1111/j.1439-0469.1979.tb00699.x [dostęp 2025-03-25] (ang.).
- ↑ Jesus Lozano-Fernandez i inni, Increasing species sampling in chelicerate genomic-scale datasets provides support for monophyly of Acari and Arachnida, „Nature Communications”, 2019, DOI: 10.1038/s41467-019-10244-7 [dostęp 2025-03-25] (ang.).
- ↑ Limulidae Leach, 1819, [w:] WoRMS – World Register of Marine Species [online] [dostęp 2025-03-15] (ang.).
Bibliografia
- Czesław Jura: Bezkręgowce: podstawy morfologii funkcjonalnej, systematyki i filogenezy. Warszawa: Wydawnictwo Naukowe PWN, 2007. ISBN 978-83-01-14595-8.
- Mały słownik zoologiczny. Bezkręgowce. Warszawa: Wiedza Powszechna, 1984. ISBN 83-214-0428-6.